

 I'm currently on a trip back east, primarily to give presentations at two different professional conferences. After meeting up with my wife, Brett, we drove to Chicago to attend the National Science Teachers' Association meeting. While there we conducted a workshop on producing virtual field trip e-books for iPads, and gave a presentation on WSC's paleontology teaching kits; Brett gave additional presentations on using mammal skulls to teach about trophic levels and on webquests. We then left Chicago and headed south to Chattanooga for the Geological Society of America Southeastern Section meeting (SEGSA).In addition to our presentations at SEGSA, Brett and I are running an exhibitor's booth (above) to give the participants information about the Western Science Center and to sell teaching kits and cast replicas of specimens to raise funds for the museum.Running our booth cuts into my time for attending presentations, but I did make it to several talks and quite a few posters (generally my favorite part of the meeting). The morning posters included a session on Geoscience Education, including one by Andy Heckert of Appalachian State on a student exercise to describe their hometown geology. In the same session Zoe Zeszut and coauthors presented on Ohio University's Digital Atlas of Ordovician Life, an online guide to the Ordovician fossils of Ohio and the surrounding regions, such as these highly fossiliferous beds just across the state line in Kentucky:
I'm currently on a trip back east, primarily to give presentations at two different professional conferences. After meeting up with my wife, Brett, we drove to Chicago to attend the National Science Teachers' Association meeting. While there we conducted a workshop on producing virtual field trip e-books for iPads, and gave a presentation on WSC's paleontology teaching kits; Brett gave additional presentations on using mammal skulls to teach about trophic levels and on webquests. We then left Chicago and headed south to Chattanooga for the Geological Society of America Southeastern Section meeting (SEGSA).In addition to our presentations at SEGSA, Brett and I are running an exhibitor's booth (above) to give the participants information about the Western Science Center and to sell teaching kits and cast replicas of specimens to raise funds for the museum.Running our booth cuts into my time for attending presentations, but I did make it to several talks and quite a few posters (generally my favorite part of the meeting). The morning posters included a session on Geoscience Education, including one by Andy Heckert of Appalachian State on a student exercise to describe their hometown geology. In the same session Zoe Zeszut and coauthors presented on Ohio University's Digital Atlas of Ordovician Life, an online guide to the Ordovician fossils of Ohio and the surrounding regions, such as these highly fossiliferous beds just across the state line in Kentucky: The afternoon session included most of the paleontology posters, including the first of my three presentations. Shortly before I left the Virginia Museum of Natural History (VMNH) I received a grant from the National Geographic Society to run a salvage operation at the Solite Quarry, a Triassic Period deposit on the Virginia/North Carolina state line that includes spectacular fossils of reptiles, fish, plants, and insects. My poster included five coauthors from VMNH, Appalachian State, and the National Museum of Scotland and was basically a progress report on the state of the salvage operation. I got the salvage operation started as one of my last activities at VMNH (below), and they continued the operation after my departure to WSC. The salvage work has resulted in the collection of a large number of new fossils, some of which were only known previously by a handful of specimens.
The afternoon session included most of the paleontology posters, including the first of my three presentations. Shortly before I left the Virginia Museum of Natural History (VMNH) I received a grant from the National Geographic Society to run a salvage operation at the Solite Quarry, a Triassic Period deposit on the Virginia/North Carolina state line that includes spectacular fossils of reptiles, fish, plants, and insects. My poster included five coauthors from VMNH, Appalachian State, and the National Museum of Scotland and was basically a progress report on the state of the salvage operation. I got the salvage operation started as one of my last activities at VMNH (below), and they continued the operation after my departure to WSC. The salvage work has resulted in the collection of a large number of new fossils, some of which were only known previously by a handful of specimens. Just to prove that I do, in fact, do field work!In some of the other paleontology section posters, Jennifer Bradham, Larisa DeSantis, and Malu Jorge used stable isotope data nd dental microwear to compare the diets of Florida specimens of the Pliocene peccaries Mylohyus (below, at top, from the Texas Memorial Museum) and Platygonus (below, at bottom, from the American Museum of Natural History):
Just to prove that I do, in fact, do field work!In some of the other paleontology section posters, Jennifer Bradham, Larisa DeSantis, and Malu Jorge used stable isotope data nd dental microwear to compare the diets of Florida specimens of the Pliocene peccaries Mylohyus (below, at top, from the Texas Memorial Museum) and Platygonus (below, at bottom, from the American Museum of Natural History):
 Three different posters examine various aspects of drill holes on mollusk shells, formed when predators drill a hole through the shell to get at the animal inside (as in the shell shown below):
Three different posters examine various aspects of drill holes on mollusk shells, formed when predators drill a hole through the shell to get at the animal inside (as in the shell shown below): Kathryn Estes-Smargiassi and three coauthors looked at these drill holes specifically on scaphopods, or tusk shells. Patricia Kelley and six coauthors examined the paleoecological patterns of drill predation on Pleistocene mollusks from Florida. And, while these drill holes are usually formed by predatory snails, Deborah Freile and three coauthors looked at examples of shells that had been drilled by octopi.Late in the afternoon I left the posters to catch the last three talks in the paleoichnology (trace fossil) session. Kelli Straka and five coauthors reported on coprolites (fossil poo) containing teeth, fish scales, and bone fragments from Triassic deposits in Arizona, while David Schwimmer presented on Cretaceous coprolites from the southeast, including examples from fish, sharks, and crocodilians, including a shark coprolite that contained almost the entire neck of a baby turtle. In the last talk in the session Tony Martin and three coauthors reported on modern alligator burrows from Georgia barrier islands, large structures that can be up to several meters long.These meeting are also a nice opportunity to keep in contact with old colleagues. I was pleasantly surprised when the WSC booth was visited by Heyo van Iten from Hanover College (below). Heyo was my undergraduate paleontology professor at Carleton College more than 25 years ago.
Kathryn Estes-Smargiassi and three coauthors looked at these drill holes specifically on scaphopods, or tusk shells. Patricia Kelley and six coauthors examined the paleoecological patterns of drill predation on Pleistocene mollusks from Florida. And, while these drill holes are usually formed by predatory snails, Deborah Freile and three coauthors looked at examples of shells that had been drilled by octopi.Late in the afternoon I left the posters to catch the last three talks in the paleoichnology (trace fossil) session. Kelli Straka and five coauthors reported on coprolites (fossil poo) containing teeth, fish scales, and bone fragments from Triassic deposits in Arizona, while David Schwimmer presented on Cretaceous coprolites from the southeast, including examples from fish, sharks, and crocodilians, including a shark coprolite that contained almost the entire neck of a baby turtle. In the last talk in the session Tony Martin and three coauthors reported on modern alligator burrows from Georgia barrier islands, large structures that can be up to several meters long.These meeting are also a nice opportunity to keep in contact with old colleagues. I was pleasantly surprised when the WSC booth was visited by Heyo van Iten from Hanover College (below). Heyo was my undergraduate paleontology professor at Carleton College more than 25 years ago. The meeting continues tomorrow with another full slate of posters and talks.
The meeting continues tomorrow with another full slate of posters and talks.
Fossil Friday - bison lower jaw
 I'm in Chicago this week for the National Science Teachers' Association meeting, in part so that my wife Brett and I can demonstrate teaching kits that we're developing based on the WSC collections (I'll have more about those in a future post). One of these teaching kits involves fossil bison, including the specimen featured in today's Fossil Friday. This is a partial right lower jaw from a bison, seen above in lateral (side) view. Below is the same specimen from above (dorsal view):
I'm in Chicago this week for the National Science Teachers' Association meeting, in part so that my wife Brett and I can demonstrate teaching kits that we're developing based on the WSC collections (I'll have more about those in a future post). One of these teaching kits involves fossil bison, including the specimen featured in today's Fossil Friday. This is a partial right lower jaw from a bison, seen above in lateral (side) view. Below is the same specimen from above (dorsal view):
 While most of this jaw is missing, it does preserve the tooth sockets ("alveolae"), even though only a single tooth is preserved. That tooth is the last molar, m3, and it's pretty unusual in occlusal view:
While most of this jaw is missing, it does preserve the tooth sockets ("alveolae"), even though only a single tooth is preserved. That tooth is the last molar, m3, and it's pretty unusual in occlusal view:
 The tooth is almost completely worn away. None of the usual complex enamel patterns that we see in bison teeth are present, because the enamel is mostly gone, except for a few patches on the edges of the tooth. Modern bison live to about 15 years in the wild, and this one had to be getting up there.But there's actually more to look at here. It's not that unusual to have a jaw fragment with only one tooth preserved; once a jaw is broken it's easy for the teeth to fall out. But if we look at the empty tooth sockets in this jaw, we can see that's not what happened here:
The tooth is almost completely worn away. None of the usual complex enamel patterns that we see in bison teeth are present, because the enamel is mostly gone, except for a few patches on the edges of the tooth. Modern bison live to about 15 years in the wild, and this one had to be getting up there.But there's actually more to look at here. It's not that unusual to have a jaw fragment with only one tooth preserved; once a jaw is broken it's easy for the teeth to fall out. But if we look at the empty tooth sockets in this jaw, we can see that's not what happened here:
 These sockets actually have bone covering the openings. That means that the teeth fell out before the animal died, and the sockets healed over! This bison was shockingly old; it had actually lived long enough for every tooth except the last one wear all the way down to the roots and fall out, with the sockets healing over. Even the last tooth was close to falling out when the animal died.It's tough to estimate the age of such an old animal, but this had to be a particularly elderly individual, and has resulted in us nicknaming it "Methuselah".
These sockets actually have bone covering the openings. That means that the teeth fell out before the animal died, and the sockets healed over! This bison was shockingly old; it had actually lived long enough for every tooth except the last one wear all the way down to the roots and fall out, with the sockets healing over. Even the last tooth was close to falling out when the animal died.It's tough to estimate the age of such an old animal, but this had to be a particularly elderly individual, and has resulted in us nicknaming it "Methuselah".
On animal cognition
 Inspired by Roy Mason's lecture at WSC last Thursday I decided to republish this post from my old blog. This was originally published at my old blog, "Updates from the Paleontology Lab" on April 6, 2011 under the title "And now for something a little different...". It seems that one of my dogs, Molly, doesn’t like large animals. I’m not sure why this should be the case, since she’s pretty much never been around any large animals except the deer that occasionally wander into our backyard. But if she spots a deer, horse, or some other large animal in the distance it generally results in a fit of barking and growling that is completely different from her normal demeanor.This behavior might seem a little out of character for Molly, but it’s not really surprising as it seems to be pretty commonplace in domestic dogs. But what I did find surprising was that Molly went into her aggressive, anti-large animal mode when she saw this:
Inspired by Roy Mason's lecture at WSC last Thursday I decided to republish this post from my old blog. This was originally published at my old blog, "Updates from the Paleontology Lab" on April 6, 2011 under the title "And now for something a little different...". It seems that one of my dogs, Molly, doesn’t like large animals. I’m not sure why this should be the case, since she’s pretty much never been around any large animals except the deer that occasionally wander into our backyard. But if she spots a deer, horse, or some other large animal in the distance it generally results in a fit of barking and growling that is completely different from her normal demeanor.This behavior might seem a little out of character for Molly, but it’s not really surprising as it seems to be pretty commonplace in domestic dogs. But what I did find surprising was that Molly went into her aggressive, anti-large animal mode when she saw this:
 Now, when we look at this we immediately think “cow”, and apparently so does Molly (or at least whatever the dog word is for “cow”). But if you really look at this, it's pretty stylized for a cow. While we think “cow” we know it’s not a real cow. But how do we know? What features are we looking at that tell us “fake cow” instead of “real cow”? Maybe it’s the wire frame, or the incorrect texture, or these and other things besides. So maybe our subconscious thought process goes something like this (and I’m just generalizing here, so don’t nitpick too much):head + ears + long body + four legs + tufted tail + cow size = cow(cow) + wire frame + non-furry texture = fake cowWhy is this interesting? Because Molly clearly wasn’t getting the “fake cow” message. She sees some feature or combination of features and thinks “cow” and ignores the features we see that tell us “fake cow”. Dogs have good eyesight, so Molly should be able to see things like the wire frame; she just doesn’t recognize the significance of those things. So her brain is taking in a subset of the data available to it, and using that subset to identify “cow”.When Molly initially saw the cow, she was in the car with the windows up. As we pulled up next to the cow, we rolled down the window, and she sniffed. Then she stopped barking and pretty much ignored the cow. So Molly’s identification process may go like this:head + ears + long body + four legs + tufted tail + cow size = cow(cow) + non-cow smell = fake cowBut she didn’t rely on the smell to make her initial identification, so presumably either of these pathways work for her:head + ears + long body + four legs + tufted tail + cow size = cowhead + ears + long body + four legs + tufted tail + cow size + cow smell = cowIt’s also interesting that the absence of a cow smell (and presence of whatever she did smell) was enough to override all her prior interpretations based on what she could see. She seems to be weighting some observations more heavily than others.In fact, her whole process seems to have a familiar pattern:Hypothesis: Based on visual observations, I think that’s a cow. If it is a cow, it should smell like a cow. (Null hypothesis: It’s not a cow.)Procedure: Sniff to see if it smells like a cow.Result: It doesn’t smell like a cow.Conclusion: It’s not a cow (the null hypothesis is not rejected).My dog is a scientist!There is one more interesting observation concerning Molly; the cow is not the first time she’s had this kind of reaction to a life-size animal model. A few years ago, she showed the same reaction to this:
Now, when we look at this we immediately think “cow”, and apparently so does Molly (or at least whatever the dog word is for “cow”). But if you really look at this, it's pretty stylized for a cow. While we think “cow” we know it’s not a real cow. But how do we know? What features are we looking at that tell us “fake cow” instead of “real cow”? Maybe it’s the wire frame, or the incorrect texture, or these and other things besides. So maybe our subconscious thought process goes something like this (and I’m just generalizing here, so don’t nitpick too much):head + ears + long body + four legs + tufted tail + cow size = cow(cow) + wire frame + non-furry texture = fake cowWhy is this interesting? Because Molly clearly wasn’t getting the “fake cow” message. She sees some feature or combination of features and thinks “cow” and ignores the features we see that tell us “fake cow”. Dogs have good eyesight, so Molly should be able to see things like the wire frame; she just doesn’t recognize the significance of those things. So her brain is taking in a subset of the data available to it, and using that subset to identify “cow”.When Molly initially saw the cow, she was in the car with the windows up. As we pulled up next to the cow, we rolled down the window, and she sniffed. Then she stopped barking and pretty much ignored the cow. So Molly’s identification process may go like this:head + ears + long body + four legs + tufted tail + cow size = cow(cow) + non-cow smell = fake cowBut she didn’t rely on the smell to make her initial identification, so presumably either of these pathways work for her:head + ears + long body + four legs + tufted tail + cow size = cowhead + ears + long body + four legs + tufted tail + cow size + cow smell = cowIt’s also interesting that the absence of a cow smell (and presence of whatever she did smell) was enough to override all her prior interpretations based on what she could see. She seems to be weighting some observations more heavily than others.In fact, her whole process seems to have a familiar pattern:Hypothesis: Based on visual observations, I think that’s a cow. If it is a cow, it should smell like a cow. (Null hypothesis: It’s not a cow.)Procedure: Sniff to see if it smells like a cow.Result: It doesn’t smell like a cow.Conclusion: It’s not a cow (the null hypothesis is not rejected).My dog is a scientist!There is one more interesting observation concerning Molly; the cow is not the first time she’s had this kind of reaction to a life-size animal model. A few years ago, she showed the same reaction to this:
 I very seriously doubt that there’s a dog word for Kentrosaurus. And yet something that Molly sees here triggers at least a “big animal” reaction, and it overrides all the (to her) unfamiliar features of a dinosaur. So what are the critical features that tell Molly that this is an animal? I have no idea, but it would be interesting to find out.OK, I admit this really doesn’t have much to do with paleontology or geology. But it’s still intriguing to think about the implications of observations like this for cognition and sensory perception.
I very seriously doubt that there’s a dog word for Kentrosaurus. And yet something that Molly sees here triggers at least a “big animal” reaction, and it overrides all the (to her) unfamiliar features of a dinosaur. So what are the critical features that tell Molly that this is an animal? I have no idea, but it would be interesting to find out.OK, I admit this really doesn’t have much to do with paleontology or geology. But it’s still intriguing to think about the implications of observations like this for cognition and sensory perception.
Fossil Friday - horse pelvis
 For this week's Fossil Friday we have the pelvic bones from a horse, collected from the Pleistocene deposits at the east end of Diamond Valley Lake.In most tetrapods (four-legged vertebrates that mostly live on land, and their descendants) the pelvic girdle is the primary structure for supporting the animal's weight and allowing it to move. As such this is usually a large complex structure made up of six separate bones (three on each side) plus several specialized, fused vertebrae (the sacrals). The pelvic bones are the ilium, the ischium, and the pubis, and in mammals these are usually fused to each other at a young age; the fused group of three bones is called the innominate.The field jacket shown above actually has portions of both the left and right innominates of a horse. The bone on the right side of the image is the left innominate, seen in dorsal view (from above), while the one on the left is the right innominate, seen in ventral view (from below). Below is a color-coded version of the same image:
For this week's Fossil Friday we have the pelvic bones from a horse, collected from the Pleistocene deposits at the east end of Diamond Valley Lake.In most tetrapods (four-legged vertebrates that mostly live on land, and their descendants) the pelvic girdle is the primary structure for supporting the animal's weight and allowing it to move. As such this is usually a large complex structure made up of six separate bones (three on each side) plus several specialized, fused vertebrae (the sacrals). The pelvic bones are the ilium, the ischium, and the pubis, and in mammals these are usually fused to each other at a young age; the fused group of three bones is called the innominate.The field jacket shown above actually has portions of both the left and right innominates of a horse. The bone on the right side of the image is the left innominate, seen in dorsal view (from above), while the one on the left is the right innominate, seen in ventral view (from below). Below is a color-coded version of the same image:
 The bones outlined in blue are the ilia; he right ilium is damaged but the left one is largely intact. The yellow outlines mark the ischia, and the red is the pubis. Only the right pubis is visible; the left one is either hidden underneath the ilium and ischium or (more likely) not preserved at all.Where the three bones come together they form a cavity called the acetabulum, outlined in green. This is the "socket" part of the hip's "ball-and-socket" joint where the leg attaches to the hip. There is also a large gap between the pubis and the ischium called the obturator foramen, which serves as a passageway for nerves and blood vessels that run through the pelvis.While there is a lot of variation is the details, all mammals that have back legs share this same basic hip structure, and even mammals that lack back legs (such as whales and sea cows) had ancestors with similar pelvic bones.
The bones outlined in blue are the ilia; he right ilium is damaged but the left one is largely intact. The yellow outlines mark the ischia, and the red is the pubis. Only the right pubis is visible; the left one is either hidden underneath the ilium and ischium or (more likely) not preserved at all.Where the three bones come together they form a cavity called the acetabulum, outlined in green. This is the "socket" part of the hip's "ball-and-socket" joint where the leg attaches to the hip. There is also a large gap between the pubis and the ischium called the obturator foramen, which serves as a passageway for nerves and blood vessels that run through the pelvis.While there is a lot of variation is the details, all mammals that have back legs share this same basic hip structure, and even mammals that lack back legs (such as whales and sea cows) had ancestors with similar pelvic bones.
Fossil Friday - juvenile mastodon femur
 This week's Fossil Friday specimen is a femur (thigh bone) from a mastodon, collected from the West Dam of Diamond Valley Lake.Like many of the Western Science Center specimens, this femur is only partially prepared, and still sits in its field jacket. The posterior side of the bone is shown above, with the back of the knee joint visible on the left. As this is the right femur, there should be a large ball joint (the femoral head) in the upper right, that would articulate with the hip socket. Unfortunately the femoral head was broken off and not preserved.The preserved part of this bone is roughly 70 cm long. That's actually pretty small for a mastodon, large specimens of which can have femora more than a meter in length. This was probably a sub-adult mastodon, although the partial fusion of the knee joint to the rest of the bone suggests that it wasn't too young.
This week's Fossil Friday specimen is a femur (thigh bone) from a mastodon, collected from the West Dam of Diamond Valley Lake.Like many of the Western Science Center specimens, this femur is only partially prepared, and still sits in its field jacket. The posterior side of the bone is shown above, with the back of the knee joint visible on the left. As this is the right femur, there should be a large ball joint (the femoral head) in the upper right, that would articulate with the hip socket. Unfortunately the femoral head was broken off and not preserved.The preserved part of this bone is roughly 70 cm long. That's actually pretty small for a mastodon, large specimens of which can have femora more than a meter in length. This was probably a sub-adult mastodon, although the partial fusion of the knee joint to the rest of the bone suggests that it wasn't too young.
Fossil Friday - amber snails
 Worldwide, easily the most common fossils are from marine invertebrates. Most sedimentary rocks are formed in the ocean, invertebrates are present in vast numbers, and the ocean is an excellent place to be buried in the mud, increasing the likelihood of being preserved. In contrast, the fossil collections at the Western Science Center are dominated by terrestrial deposits from Riverside County, so most of our fossils are land vertebrates. We do, however, have a pretty good collection of terrestrial and freshwater invertebrates in the collections.Above is the shell of an amber snail (Gastropoda) from the genus Succinea. Amber snails are air-breathing snails (they have lungs) that generally live in marshy areas. The genus is wide-ranging, and modern Succinea are found all over North America. Below is the same shell in a different orientation, showing the aperture (the opening to the inside of the shell):
Worldwide, easily the most common fossils are from marine invertebrates. Most sedimentary rocks are formed in the ocean, invertebrates are present in vast numbers, and the ocean is an excellent place to be buried in the mud, increasing the likelihood of being preserved. In contrast, the fossil collections at the Western Science Center are dominated by terrestrial deposits from Riverside County, so most of our fossils are land vertebrates. We do, however, have a pretty good collection of terrestrial and freshwater invertebrates in the collections.Above is the shell of an amber snail (Gastropoda) from the genus Succinea. Amber snails are air-breathing snails (they have lungs) that generally live in marshy areas. The genus is wide-ranging, and modern Succinea are found all over North America. Below is the same shell in a different orientation, showing the aperture (the opening to the inside of the shell):
 Succinea is a tiny snail; the black box the shell is sitting on in the images is only 1 cm across! A fairly large number of Succinea shells were recovered in the Pleistocene deposits from Diamond Valley Lake.
Succinea is a tiny snail; the black box the shell is sitting on in the images is only 1 cm across! A fairly large number of Succinea shells were recovered in the Pleistocene deposits from Diamond Valley Lake.
Fossil Friday - Bison antiquus skull
 Bison are among the most common large animals from the Diamond Valley Lake region, and we have a number of nice specimens at the Western Science Center. Shown above is a cranium of Bison antiquus, one of several such skulls in our collection.This skull, while incomplete, is in pretty good shape. It's still in its field jacket, and only partially prepared, so only the ventral (bottom) side is visible. The front of the skull is to the left. The left horn core is noticeably missing, and there is some crushing of the skull that's most clearly visible along the right side (bottom of the image). The 2nd premolar is missing on each side, but the other teeth are all present. If you're looking carefully, you might have noticed that there are no incisor teeth in the front of the mouth. Bison don't have incisors in the upper jaws, a character they share with other "horned ruminants" (Infraorder Pecora). The mass of bone at the back of the skull (on the right side of the image) is the atlas (first neck vertebra), more-or-less still attached to the skull.There are two different species of bison present in the Pleistocene deposits at Diamond Valley Lake, the smaller Bison antiquus and the larger, longer-horned Bison latifrons. (When I say "smaller", that's relative; B. antiquus was still larger than modern bison.) Bison antiquus seems to be the more common of the two fossil species in this area. It's generally thought that B. antiquus is the direct ancestor to the modern bison, Bison bison.
Bison are among the most common large animals from the Diamond Valley Lake region, and we have a number of nice specimens at the Western Science Center. Shown above is a cranium of Bison antiquus, one of several such skulls in our collection.This skull, while incomplete, is in pretty good shape. It's still in its field jacket, and only partially prepared, so only the ventral (bottom) side is visible. The front of the skull is to the left. The left horn core is noticeably missing, and there is some crushing of the skull that's most clearly visible along the right side (bottom of the image). The 2nd premolar is missing on each side, but the other teeth are all present. If you're looking carefully, you might have noticed that there are no incisor teeth in the front of the mouth. Bison don't have incisors in the upper jaws, a character they share with other "horned ruminants" (Infraorder Pecora). The mass of bone at the back of the skull (on the right side of the image) is the atlas (first neck vertebra), more-or-less still attached to the skull.There are two different species of bison present in the Pleistocene deposits at Diamond Valley Lake, the smaller Bison antiquus and the larger, longer-horned Bison latifrons. (When I say "smaller", that's relative; B. antiquus was still larger than modern bison.) Bison antiquus seems to be the more common of the two fossil species in this area. It's generally thought that B. antiquus is the direct ancestor to the modern bison, Bison bison.
Darwin's other theory (repost)
 Diagrams from Darwin, 1842.I originally published this on my old blog, Updates from the Paleontology Lab, on March 24, 2010. I'm republishing it here for Darwin Day.Tim has to write an essay about a famous scientist for his science class that includes describing that person’s major contributions, and he chose Charles Darwin (that’s my boy!). To help him out, I showed him some of my Keynote presentations from my historical geology classes (I’m not teaching this semester, but I still have all my lecture slides). I came across some slides that I thought were worth reproducing here.Darwin is obviously most famous for his work on evolution and the publication of “Origin of Species” in 1859, but that wasn’t all he did in his career. Among geologists, he’s also well known for his theories on coral reef development, which like evolution largely stemmed from his travels on the HMS Beagle. These thoughts were published in “The Structure and Distribution of Coral Reefs” in 1842 (available online here).Darwin started by describing in detail coral reefs all over the world, concentrating on those in the Pacific and Indian Oceans that he visited while on the Beagle (see map at top). His Plate 1 included maps of some of these islands, modified from excellent Royal Navy maps:
Diagrams from Darwin, 1842.I originally published this on my old blog, Updates from the Paleontology Lab, on March 24, 2010. I'm republishing it here for Darwin Day.Tim has to write an essay about a famous scientist for his science class that includes describing that person’s major contributions, and he chose Charles Darwin (that’s my boy!). To help him out, I showed him some of my Keynote presentations from my historical geology classes (I’m not teaching this semester, but I still have all my lecture slides). I came across some slides that I thought were worth reproducing here.Darwin is obviously most famous for his work on evolution and the publication of “Origin of Species” in 1859, but that wasn’t all he did in his career. Among geologists, he’s also well known for his theories on coral reef development, which like evolution largely stemmed from his travels on the HMS Beagle. These thoughts were published in “The Structure and Distribution of Coral Reefs” in 1842 (available online here).Darwin started by describing in detail coral reefs all over the world, concentrating on those in the Pacific and Indian Oceans that he visited while on the Beagle (see map at top). His Plate 1 included maps of some of these islands, modified from excellent Royal Navy maps:
 Compare to modern Google Earth images of the same islands:
Compare to modern Google Earth images of the same islands:
 Darwin pointed out that there were three distinct types of reefs: fringing reefs which were right next to an island’s shore (Kosrae, below), barrier reefs which surrounded an island and were separated from it by a lagoon (Bora bora), and atolls, ring-shaped reefs surrounding a lagoon with no central island (Hao).
Darwin pointed out that there were three distinct types of reefs: fringing reefs which were right next to an island’s shore (Kosrae, below), barrier reefs which surrounded an island and were separated from it by a lagoon (Bora bora), and atolls, ring-shaped reefs surrounding a lagoon with no central island (Hao).
 (As an aside, Darwin apparently established the regular use of the term atoll in the western world, even though it had been used by some earlier European explorers. From page 2: “As the reef of a lagoon-island generally supports many separate small islands, the word ‘island,’ applied to the whole, is often the cause of confusion; hence I have invariably used in this volume the term ‘atoll,’ which is the name given to these circular groups of coral islets by their inhabitants in the Indian Ocean, and is synonymous with ‘lagoon-island.’)With his usual meticulous attention to detail, he went through numerous examples of each type of reef, looking at how the coral were distributed laterally and vertically, what kinds of rocks were found in the area, and so on. He pulled up samples from various depths of water to show that coral couldn’t live at depth greater than about 200 feet, even though dead corals were found much deeper than that. (His descriptions include this remarkable quote on page 80: “A little further out the depth is thirty fathoms, and thence the bank slopes rapidly into the depths of the ocean. … The water was so clear outside the reef, that I could distinguish every object forming the rugged bottom.”)Based on these observations he proposed that these three types of reef are all related to each other. Essentially atolls are formed when an island with a fringing reef subsides, but the coral growth keeps pace with the subsidence. Therefore, atolls had encountered greater subsidence than fringing reefs, and barrier reefs were intermediate. Here are Darwin’s cross-section interpretations:
(As an aside, Darwin apparently established the regular use of the term atoll in the western world, even though it had been used by some earlier European explorers. From page 2: “As the reef of a lagoon-island generally supports many separate small islands, the word ‘island,’ applied to the whole, is often the cause of confusion; hence I have invariably used in this volume the term ‘atoll,’ which is the name given to these circular groups of coral islets by their inhabitants in the Indian Ocean, and is synonymous with ‘lagoon-island.’)With his usual meticulous attention to detail, he went through numerous examples of each type of reef, looking at how the coral were distributed laterally and vertically, what kinds of rocks were found in the area, and so on. He pulled up samples from various depths of water to show that coral couldn’t live at depth greater than about 200 feet, even though dead corals were found much deeper than that. (His descriptions include this remarkable quote on page 80: “A little further out the depth is thirty fathoms, and thence the bank slopes rapidly into the depths of the ocean. … The water was so clear outside the reef, that I could distinguish every object forming the rugged bottom.”)Based on these observations he proposed that these three types of reef are all related to each other. Essentially atolls are formed when an island with a fringing reef subsides, but the coral growth keeps pace with the subsidence. Therefore, atolls had encountered greater subsidence than fringing reefs, and barrier reefs were intermediate. Here are Darwin’s cross-section interpretations:
 Darwin pretty much got it right. The one thing he missed (and no one realized it for more than 100 years) concerned the nature of the subsidence that formed the atolls. Darwin noticed that the atolls tended to cluster in certain areas, and believed those areas were subject to greater rates of subsidence. In fact, the atolls had encountered more subsidence not because the rate of subsidence was particularly higher there, but because they were older than the other islands.It finally took the development of plate tectonics theory in the 1960’s to fully understand Darwin’s observations. Any given island chain (such as Hawaii) is formed from a single volcanic hot spot. As the Pacific plate moves over the hot spot, the old volcanic island is carried away and a new one forms, with a new fringing reef surrounding it. In the Hawaiian chain, Midway and Oahu are probably experiencing close to the same subsidence rates. But Midway (an atoll) has been experiencing that subsidence for about 27 million years, while Oahu (with a fringing reef) has only been subsiding for about 3 million years.While Darwin played the key role in the development of the most important unifying theory in biological sciences (evolution), he also played a small but significant role in the development of the major unifying theory in geological sciences (plate tectonics).Reference:Darwin, C., 1942. The Structure and Distribution of Coral Reefs. Smith, Elder, and Co., London, 214 p.
Darwin pretty much got it right. The one thing he missed (and no one realized it for more than 100 years) concerned the nature of the subsidence that formed the atolls. Darwin noticed that the atolls tended to cluster in certain areas, and believed those areas were subject to greater rates of subsidence. In fact, the atolls had encountered more subsidence not because the rate of subsidence was particularly higher there, but because they were older than the other islands.It finally took the development of plate tectonics theory in the 1960’s to fully understand Darwin’s observations. Any given island chain (such as Hawaii) is formed from a single volcanic hot spot. As the Pacific plate moves over the hot spot, the old volcanic island is carried away and a new one forms, with a new fringing reef surrounding it. In the Hawaiian chain, Midway and Oahu are probably experiencing close to the same subsidence rates. But Midway (an atoll) has been experiencing that subsidence for about 27 million years, while Oahu (with a fringing reef) has only been subsiding for about 3 million years.While Darwin played the key role in the development of the most important unifying theory in biological sciences (evolution), he also played a small but significant role in the development of the major unifying theory in geological sciences (plate tectonics).Reference:Darwin, C., 1942. The Structure and Distribution of Coral Reefs. Smith, Elder, and Co., London, 214 p.
Fossil Friday - horse thoracic vertebrae
 Our Fossil Friday specimen for this week is a pair of Pleistocene horse vertebrae, still in their field jacket.These two vertebrae are the sixth and seventh thoracics (see this link for an overview of vertebral regions in mammals). In the image the bottoms of the vertebrae are to the left and the tops are to the right. The sixth thoracic is the upper of the two, and you're looking at the posterior surface. In contrast, anterior surface of the seventh thoracic is visible. In the articulated skeleton these vertebrae lie roughly adjacent to the shoulder blades, as shown below in this mounted skeleton at the Texas Memorial Museum:
Our Fossil Friday specimen for this week is a pair of Pleistocene horse vertebrae, still in their field jacket.These two vertebrae are the sixth and seventh thoracics (see this link for an overview of vertebral regions in mammals). In the image the bottoms of the vertebrae are to the left and the tops are to the right. The sixth thoracic is the upper of the two, and you're looking at the posterior surface. In contrast, anterior surface of the seventh thoracic is visible. In the articulated skeleton these vertebrae lie roughly adjacent to the shoulder blades, as shown below in this mounted skeleton at the Texas Memorial Museum:
 The long projections coming from the top of each vertebra are called "neural spines" or "spinous processes". They are rather tall in horses, especially in the anterior half of the thoracic region. These tall processes form the horse's withers, the hump present above the shoulders:
The long projections coming from the top of each vertebra are called "neural spines" or "spinous processes". They are rather tall in horses, especially in the anterior half of the thoracic region. These tall processes form the horse's withers, the hump present above the shoulders:
 Tall neural spines are present in lots of different animals, and can serve a variety of functions including display and supporting fat bodies. They're pretty much always present in land mammals like horses that have large, heavy heads. Horses spend the vast majority of their time in a posture like the one in the image above; head to the ground, eating. It's hard to keep balanced with that heavy head stuck out like that, and ligaments that run along the neural spines transfer much of the load throughout the vertebral column. The neural spines also serve as attachment points for muscles that move and support the head, neck, and shoulders.These two vertebrae were collected in 1997 near the east dam of Diamond Valley Lake.
Tall neural spines are present in lots of different animals, and can serve a variety of functions including display and supporting fat bodies. They're pretty much always present in land mammals like horses that have large, heavy heads. Horses spend the vast majority of their time in a posture like the one in the image above; head to the ground, eating. It's hard to keep balanced with that heavy head stuck out like that, and ligaments that run along the neural spines transfer much of the load throughout the vertebral column. The neural spines also serve as attachment points for muscles that move and support the head, neck, and shoulders.These two vertebrae were collected in 1997 near the east dam of Diamond Valley Lake.
Fossil Friday - baby mastodon teeth
 It seems that most of the mastodon remains from Diamond Valley Lake are from adult or nearly adult animals, although there are exceptions representing younger animals. Then we have the example shown here, from an almost ridiculously cute baby mastodon.This tooth is the upper right second premolar (dP2), the very first upper tooth to erupt in mastodons. It's shown above in occlusal view, with the front of the tooth to the left. Mastodon teeth have distinctive transverse ridges of enamel (they're what make tooth look "bumpy"), and the number of ridges differs by tooth position. Typically the second and third premolars each have two ridges, the fourth premolar and first two molars each have three, and the third molar has five. The two ridges in this tooth make it either a second or third premolar, but its size gives away its position; this tooth is only 31 mm long! This puts it squarely in the size range of dP2 specimens from Florida described by Green and Hulbert (2005), which ranged from 27.6 to 36.4 mm. To get an idea of just how tiny this tooth is, here it is beside a mastodon third molar:
It seems that most of the mastodon remains from Diamond Valley Lake are from adult or nearly adult animals, although there are exceptions representing younger animals. Then we have the example shown here, from an almost ridiculously cute baby mastodon.This tooth is the upper right second premolar (dP2), the very first upper tooth to erupt in mastodons. It's shown above in occlusal view, with the front of the tooth to the left. Mastodon teeth have distinctive transverse ridges of enamel (they're what make tooth look "bumpy"), and the number of ridges differs by tooth position. Typically the second and third premolars each have two ridges, the fourth premolar and first two molars each have three, and the third molar has five. The two ridges in this tooth make it either a second or third premolar, but its size gives away its position; this tooth is only 31 mm long! This puts it squarely in the size range of dP2 specimens from Florida described by Green and Hulbert (2005), which ranged from 27.6 to 36.4 mm. To get an idea of just how tiny this tooth is, here it is beside a mastodon third molar:
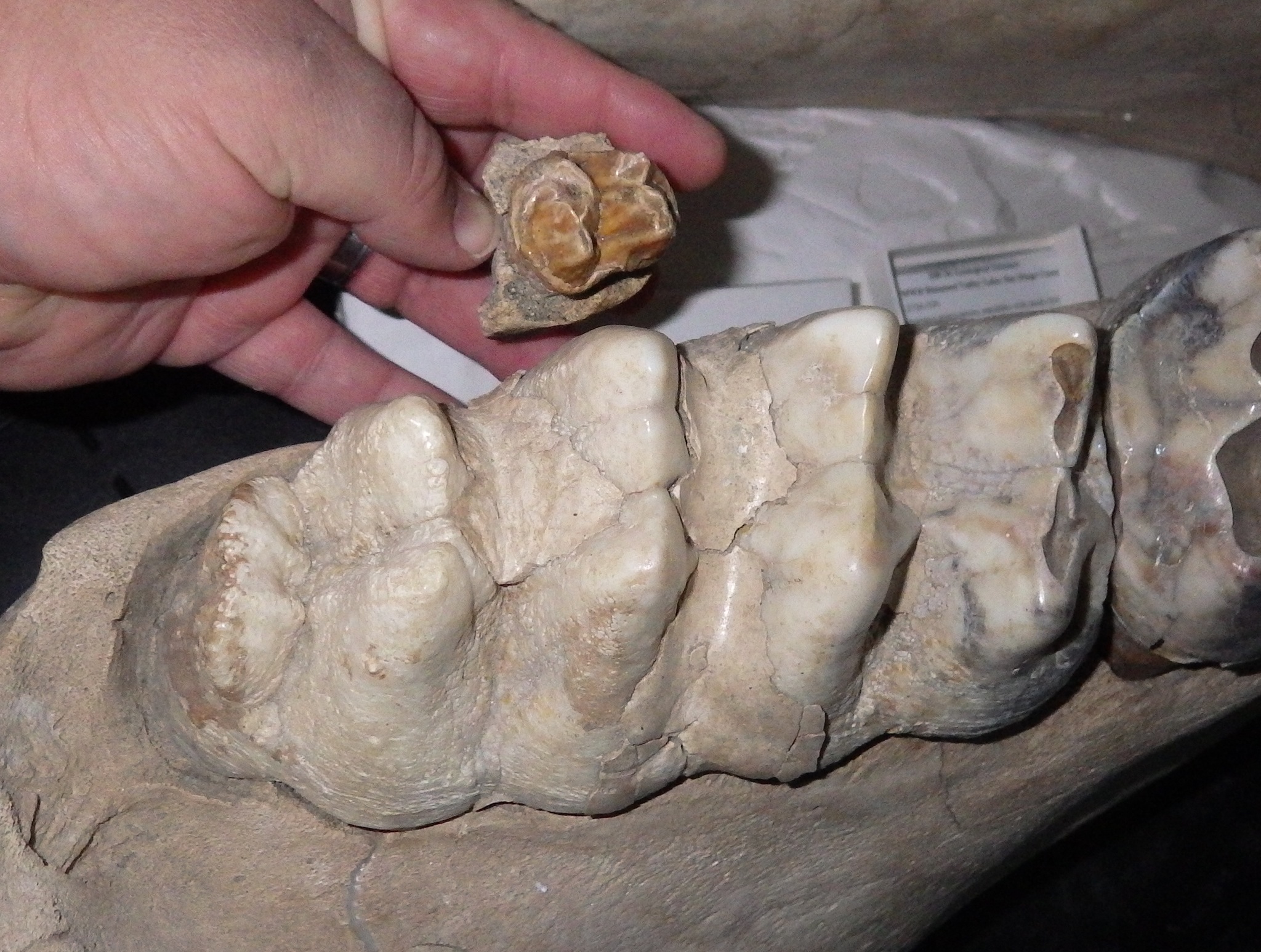 Here's a labial (side) view of the premolar:
Here's a labial (side) view of the premolar:
 This tooth was discovered near the West Dam in December 1997. It turns out that some other baby mastodon teeth were found at the same site three days earlier. While they have separate numbers in the collection, it's very likely that these teeth all came from one animal. The additional material includes the left maxilla fragment below, shown in labial view with the front toward the left:
This tooth was discovered near the West Dam in December 1997. It turns out that some other baby mastodon teeth were found at the same site three days earlier. While they have separate numbers in the collection, it's very likely that these teeth all came from one animal. The additional material includes the left maxilla fragment below, shown in labial view with the front toward the left:
 Here's the same fragment in occlusal view, again with the front toward the left:
Here's the same fragment in occlusal view, again with the front toward the left:
 The larger, complete tooth is the fourth premolar (note the three transverse enamel ridges). In front of it is the posterior ridge of the third premolar.The fragmentary right fourth premolar was also collected (occlusal view, anterior to the right):
The larger, complete tooth is the fourth premolar (note the three transverse enamel ridges). In front of it is the posterior ridge of the third premolar.The fragmentary right fourth premolar was also collected (occlusal view, anterior to the right):
 If all these pieces came from one individual (which I think is very likely), we have almost the entire upper left premolar series, lacking only the front half of the third premolar, as well as part of one right premolar.So I called this a baby mastodon, but how young was it? We can estimate the age if we assume that mastodon growth rates and tooth eruption times were similar to those of modern elephants. Mastodonts and elephants are only distantly related, so undoubtedly this introduces some uncertainty, but given their similar sizes and tooth replacement patterns it should produce a reasonable estimate.A key feature to look at is the amount of wear on each tooth. The fourth premolars and the preserved part of the third premolar show no wear at all. But in the second premolar, while the enamel ridges are mostly intact, there is a little wear at the tips (this is most clearly visible in the occlusal view at the top). In African elephants, the second premolar begins to wear when the elephant is about 2 months old, and by the age of 2 years the tooth has worn down and fallen out. Given the light wear present on this tooth, this mastodon was likely no more than 4 to 6 months old!Reference: Green, J. L. And R. C. Hulbert, Jr. 2005. The deciduous premolars of Mammut americanum (Mammalia, Proboscidea). Journal of Vertebrate Paleontology 25:702-715.
If all these pieces came from one individual (which I think is very likely), we have almost the entire upper left premolar series, lacking only the front half of the third premolar, as well as part of one right premolar.So I called this a baby mastodon, but how young was it? We can estimate the age if we assume that mastodon growth rates and tooth eruption times were similar to those of modern elephants. Mastodonts and elephants are only distantly related, so undoubtedly this introduces some uncertainty, but given their similar sizes and tooth replacement patterns it should produce a reasonable estimate.A key feature to look at is the amount of wear on each tooth. The fourth premolars and the preserved part of the third premolar show no wear at all. But in the second premolar, while the enamel ridges are mostly intact, there is a little wear at the tips (this is most clearly visible in the occlusal view at the top). In African elephants, the second premolar begins to wear when the elephant is about 2 months old, and by the age of 2 years the tooth has worn down and fallen out. Given the light wear present on this tooth, this mastodon was likely no more than 4 to 6 months old!Reference: Green, J. L. And R. C. Hulbert, Jr. 2005. The deciduous premolars of Mammut americanum (Mammalia, Proboscidea). Journal of Vertebrate Paleontology 25:702-715.
Fossil Friday - pocket gopher skull
 While the Pleistocene deposits from Diamond Valley Lake have lots of big impressive animals, small animals are actually much more numerous. The most common mammal in "The Valley of the Mastodons" is the pocket gopher!While most of our pocket gopher (genus Thomomys) remains are isolated teeth, there are a few more substantial specimens including the partial cranium shown here. At top is the skull in dorsal (top) view, with the front of the skull to the left. Most of the back and left side of the skull is missing, but the front and part of the right side are in pretty good shape. The ring of bone on the right side of the skull (at the top of the image) is made up of mostly of the jugal and the zygomatic process of the squamosal. Together these bones form the outer margin of the orbit (eye socket) and the temporal fossa, an opening for the muscles that close the lower jaw.Below is a lateral view, showing the right side of the cranium:
While the Pleistocene deposits from Diamond Valley Lake have lots of big impressive animals, small animals are actually much more numerous. The most common mammal in "The Valley of the Mastodons" is the pocket gopher!While most of our pocket gopher (genus Thomomys) remains are isolated teeth, there are a few more substantial specimens including the partial cranium shown here. At top is the skull in dorsal (top) view, with the front of the skull to the left. Most of the back and left side of the skull is missing, but the front and part of the right side are in pretty good shape. The ring of bone on the right side of the skull (at the top of the image) is made up of mostly of the jugal and the zygomatic process of the squamosal. Together these bones form the outer margin of the orbit (eye socket) and the temporal fossa, an opening for the muscles that close the lower jaw.Below is a lateral view, showing the right side of the cranium:
 The long, curved shiny feature at the front of the skull (on the right side of the photo) are the incisor teeth. The presence of a single pair of enlarged, ever-growing incisors is a feature shared among all rodents. Unlike other gophers, in Thomomys the incisors are smooth and lack longitudinal ridges (one common name for Thomomys is the "smooth toothed pocket gopher").Here is the ventral (bottom) side of the skull:
The long, curved shiny feature at the front of the skull (on the right side of the photo) are the incisor teeth. The presence of a single pair of enlarged, ever-growing incisors is a feature shared among all rodents. Unlike other gophers, in Thomomys the incisors are smooth and lack longitudinal ridges (one common name for Thomomys is the "smooth toothed pocket gopher").Here is the ventral (bottom) side of the skull:
 While there' some damage present here, and some cleaning is still needed, it looks like most of the tooth row is preserved. The complex tooth at the front of the row is the fourth premolar, while the large peg-like tooth that is partially sticking out of its socket is either the second or third molar.Pocket gophers are still common in western North America. As burrowing animals, their preservation potential is pretty high; it's a lot easier to become a fossil if you already live most of your life underground!
While there' some damage present here, and some cleaning is still needed, it looks like most of the tooth row is preserved. The complex tooth at the front of the row is the fourth premolar, while the large peg-like tooth that is partially sticking out of its socket is either the second or third molar.Pocket gophers are still common in western North America. As burrowing animals, their preservation potential is pretty high; it's a lot easier to become a fossil if you already live most of your life underground!
Fossil Friday - tusk fragment
 One of the prominent features of modern elephants are tusks, which are greatly enlarged upper incisor teeth. The presence of tusks is shared with most of the elephants' extinct proboscidean relatives, including mammoths and mastodons.Besides their size, elephant tusks are a bit different from most mammal teeth in that they lack enamel; tusks are made of ivory, which is dense dentine covered with cementum. Elephants use their tusks for a variety of tasks, including fighting, scraping bark off trees, and helping to loosen sediment when digging water holes, and the ivory tusks will get worn and polished at the tips. If the use is too vigorous the tip of the tusk may break off entirely. This isn't necessarily a problem for the elephant, because the tusks grow continuously. But it does mean that elephants may leave certain areas littered with broken tusk fragments.Mammoths and mastodons had tusks that were structurally very similar to those of modern elephants, and there they appear to have used them in similar ways. Their tusks were also just as prone to breakage, and isolated fragments often turn up as fossils. Shown above is the broken tip of a tusk that found near the western end of Diamond Valley Lake. Unfortunately, there are no morphological features present to tell if this came from a mammoth or a mastodon (at least none that I can identify). The tip (on the left) is polished and rounded from heavy wear, while the broken edge on the right is sharp and jagged. Below is a view of the other side of the same fragment:
One of the prominent features of modern elephants are tusks, which are greatly enlarged upper incisor teeth. The presence of tusks is shared with most of the elephants' extinct proboscidean relatives, including mammoths and mastodons.Besides their size, elephant tusks are a bit different from most mammal teeth in that they lack enamel; tusks are made of ivory, which is dense dentine covered with cementum. Elephants use their tusks for a variety of tasks, including fighting, scraping bark off trees, and helping to loosen sediment when digging water holes, and the ivory tusks will get worn and polished at the tips. If the use is too vigorous the tip of the tusk may break off entirely. This isn't necessarily a problem for the elephant, because the tusks grow continuously. But it does mean that elephants may leave certain areas littered with broken tusk fragments.Mammoths and mastodons had tusks that were structurally very similar to those of modern elephants, and there they appear to have used them in similar ways. Their tusks were also just as prone to breakage, and isolated fragments often turn up as fossils. Shown above is the broken tip of a tusk that found near the western end of Diamond Valley Lake. Unfortunately, there are no morphological features present to tell if this came from a mammoth or a mastodon (at least none that I can identify). The tip (on the left) is polished and rounded from heavy wear, while the broken edge on the right is sharp and jagged. Below is a view of the other side of the same fragment:
 This surface has a wrinkled appearance, and a rounded groove running for most of its length. I believe this represents an earlier break, in which the broken surface was smoothed off and polished prior to the entire tip breaking off. If that's correct, then this small fragment actually records two tusk-breakage events with a substantial amount of time between them.It's interesting to note that, according to Haynes (1991), while African elephants use their tusks for a variety of activities, they normally only break them when fighting each other. This seems to be especially prevalent drought years, when elephants will fight each other over access to water holes, although male elephants will fight in dominance displays, and there's evidence that mammoths and mastodons engaged in similar activities.Reference: Haynes, G., 1991. Mammoths, Mastodonts, and Elephants. Cambridge University Press, Cambridge, 413 p.
This surface has a wrinkled appearance, and a rounded groove running for most of its length. I believe this represents an earlier break, in which the broken surface was smoothed off and polished prior to the entire tip breaking off. If that's correct, then this small fragment actually records two tusk-breakage events with a substantial amount of time between them.It's interesting to note that, according to Haynes (1991), while African elephants use their tusks for a variety of activities, they normally only break them when fighting each other. This seems to be especially prevalent drought years, when elephants will fight each other over access to water holes, although male elephants will fight in dominance displays, and there's evidence that mammoths and mastodons engaged in similar activities.Reference: Haynes, G., 1991. Mammoths, Mastodonts, and Elephants. Cambridge University Press, Cambridge, 413 p.
Fossil Friday - bison horn
 Bison are among the most common animals in the Diamond Valley Lake Local Fauna. While no complete bison skeletons were recovered during excavations for the construction of the reservoir, large numbers of fragments, individual bones and teeth, and associated bones were discovered. Some of these are impressively large, including the horn core shown here.Bison are members of the Family Bovidae, a diverse group that includes cattle, goats, sheep, and antelope. A characteristic shared by practically all bovids is the presence of horns in males (there are some domestic bovid breeds in which the males lack horns); in many species the females have horns as well. The horns vary greatly in size and shape between species, but they always have certain features in common: they form as outgrowths of the frontal bones on the roof of the skull, they do not branch, and they are covered in a keratin sheath. Keratin doesn't usually preserve in fossils, so what we generally see in fossils is the rough bone underneath. Because the horn is part of the frontal bone, it is not shed each year like the antlers of deer.There are two different extinct species of bison known from Diamond Valley, Bison antiquus and Bison latifrons; among other differences, B. latifrons is larger and has much longer horns. As big as this specimen is, it's not immediately clear which species it represents. It's on the small side for B. latifrons, but a little larger than some of our B. antiquus specimens.
Bison are among the most common animals in the Diamond Valley Lake Local Fauna. While no complete bison skeletons were recovered during excavations for the construction of the reservoir, large numbers of fragments, individual bones and teeth, and associated bones were discovered. Some of these are impressively large, including the horn core shown here.Bison are members of the Family Bovidae, a diverse group that includes cattle, goats, sheep, and antelope. A characteristic shared by practically all bovids is the presence of horns in males (there are some domestic bovid breeds in which the males lack horns); in many species the females have horns as well. The horns vary greatly in size and shape between species, but they always have certain features in common: they form as outgrowths of the frontal bones on the roof of the skull, they do not branch, and they are covered in a keratin sheath. Keratin doesn't usually preserve in fossils, so what we generally see in fossils is the rough bone underneath. Because the horn is part of the frontal bone, it is not shed each year like the antlers of deer.There are two different extinct species of bison known from Diamond Valley, Bison antiquus and Bison latifrons; among other differences, B. latifrons is larger and has much longer horns. As big as this specimen is, it's not immediately clear which species it represents. It's on the small side for B. latifrons, but a little larger than some of our B. antiquus specimens.
Fossil Friday - mastodon premolar
 I'm starting 2015 at Valley of the Mastodon with an example of the blog's namesake, a tooth from a mastodon.This tooth is much smaller than the one I showed for Fossil Friday back in October, and represents a much younger animal. Like elephants, mastodons grow a total of six teeth in each quarter of their jaws (not including tusks), deciduous premolars 2-4 and molars 1-3. However, these teeth erupt sequentially as if they were on a conveyer belt, so there are never more than three teeth in the mouth at any one time.The tooth is shown above in occlusal view, with the front of the tooth to the right. It seems to have been rolled around in a stream or otherwise eroded, as there is a fair amount of damage including missing roots. There is also a lot of wear on the tooth caused by chewing, so that the enamel ridges characteristic of mastodons have been worn down, leaving three oval enamel ridges on the occlusal surface of the tooth. The fact that there are three of these enamel ovals suggests that this was not the 2nd or 3rd premolar, as these teeth usually only have two enamel ridges. The tooth is also small, only about 7 cm long, and is about twice as long as it is wide, suggesting that it is the 4th premolar. Moreover, the enamel ridges are not perpendicular to the long axis of the tooth, suggesting that this is a lower premolar rather than an upper one.Below is a labial view of the same tooth, showing the heavily worn crown and the damaged roots:
I'm starting 2015 at Valley of the Mastodon with an example of the blog's namesake, a tooth from a mastodon.This tooth is much smaller than the one I showed for Fossil Friday back in October, and represents a much younger animal. Like elephants, mastodons grow a total of six teeth in each quarter of their jaws (not including tusks), deciduous premolars 2-4 and molars 1-3. However, these teeth erupt sequentially as if they were on a conveyer belt, so there are never more than three teeth in the mouth at any one time.The tooth is shown above in occlusal view, with the front of the tooth to the right. It seems to have been rolled around in a stream or otherwise eroded, as there is a fair amount of damage including missing roots. There is also a lot of wear on the tooth caused by chewing, so that the enamel ridges characteristic of mastodons have been worn down, leaving three oval enamel ridges on the occlusal surface of the tooth. The fact that there are three of these enamel ovals suggests that this was not the 2nd or 3rd premolar, as these teeth usually only have two enamel ridges. The tooth is also small, only about 7 cm long, and is about twice as long as it is wide, suggesting that it is the 4th premolar. Moreover, the enamel ridges are not perpendicular to the long axis of the tooth, suggesting that this is a lower premolar rather than an upper one.Below is a labial view of the same tooth, showing the heavily worn crown and the damaged roots:
 Because of the sequential tooth replacement in mastodons and elephants, the 4th premolar is only present in the mouth for a fairly short period of time. For modern elephants this tooth is only present from about 3 to 10 years of age. Assuming mastodonts grew at the same rate as elephants, this tooth probably represents a mastodon that was roughly 10 years old.
Because of the sequential tooth replacement in mastodons and elephants, the 4th premolar is only present in the mouth for a fairly short period of time. For modern elephants this tooth is only present from about 3 to 10 years of age. Assuming mastodonts grew at the same rate as elephants, this tooth probably represents a mastodon that was roughly 10 years old.
Fossil Friday - Tyrannosaurus tooth
 The vast majority of the Western Science Center's fossil collection comes from Pleistocene deposits in Riverside County, so most of our specimens are less than 200,000 years old. Being so geologically young, birds are almost the only dinosaurs in our collection, as birds were the only dinosaurs to survive the mass extinction at the end of the Cretaceous Period 65 million years ago. We do, however, have a handful of much older, non-avian dinosaurs at the museum.On the left above is a partial single tooth from Tyrannosaurus rex from the Late Cretaceous Hell Creek Formation in Montana, which was collected by Harley Garbani and donated to the museum by Harley and his wife Mary. The tooth was apparently found isolated, which is not uncommon with dinosaurs as they shed teeth throughout their lives. As is typical of Tyrannosaurus and other theropods, the tooth has serrated cutting edges, one of which is visible near the left edge of the tooth. There is noticeable apical wear at the tip of the tooth, which is somewhat rounded off.This tooth was one of the first subjects for our molding and casting program. At the center of the photo is an unpainted resin cast of the tooth, and on the right is a painted cast. The serrated cutting edge is actually more visible in the cast than in the original specimen.The original specimen is on display at the Western Science Center as part of the "Harley Garbani: Dinosaur Hunter" exhibit. Replicas of the tooth will be available in the museum store early next year.
The vast majority of the Western Science Center's fossil collection comes from Pleistocene deposits in Riverside County, so most of our specimens are less than 200,000 years old. Being so geologically young, birds are almost the only dinosaurs in our collection, as birds were the only dinosaurs to survive the mass extinction at the end of the Cretaceous Period 65 million years ago. We do, however, have a handful of much older, non-avian dinosaurs at the museum.On the left above is a partial single tooth from Tyrannosaurus rex from the Late Cretaceous Hell Creek Formation in Montana, which was collected by Harley Garbani and donated to the museum by Harley and his wife Mary. The tooth was apparently found isolated, which is not uncommon with dinosaurs as they shed teeth throughout their lives. As is typical of Tyrannosaurus and other theropods, the tooth has serrated cutting edges, one of which is visible near the left edge of the tooth. There is noticeable apical wear at the tip of the tooth, which is somewhat rounded off.This tooth was one of the first subjects for our molding and casting program. At the center of the photo is an unpainted resin cast of the tooth, and on the right is a painted cast. The serrated cutting edge is actually more visible in the cast than in the original specimen.The original specimen is on display at the Western Science Center as part of the "Harley Garbani: Dinosaur Hunter" exhibit. Replicas of the tooth will be available in the museum store early next year.
Fossil Friday - bison molar
 As our molding and casting program gets underway, we've been going through the WSC collections looking for good candidates for molding. We've been particularly interested in teeth, because in many mammals the teeth are highly distinctive and potentially provide a lot of information about the animal. Over the last week we've been focusing on bison teeth.Shown above is a more-or-less complete lower right molar from Bison, probably the 2nd molar, shown in labial view. Here's the same tooth in lingual view:
As our molding and casting program gets underway, we've been going through the WSC collections looking for good candidates for molding. We've been particularly interested in teeth, because in many mammals the teeth are highly distinctive and potentially provide a lot of information about the animal. Over the last week we've been focusing on bison teeth.Shown above is a more-or-less complete lower right molar from Bison, probably the 2nd molar, shown in labial view. Here's the same tooth in lingual view:
 In this view there's a ridge running vertically down the middle of the tooth called a stylid. This is a column of enamel that will show up on the occlusal surface of the tooth as a small ring of enamel if the tooth is sufficiently worn. This stylid seems to always be present in Bison, but in the closely related genus Bos (cows) the stylid is usually absent or only weakly developed.Here's the tooth in occlusal view:
In this view there's a ridge running vertically down the middle of the tooth called a stylid. This is a column of enamel that will show up on the occlusal surface of the tooth as a small ring of enamel if the tooth is sufficiently worn. This stylid seems to always be present in Bison, but in the closely related genus Bos (cows) the stylid is usually absent or only weakly developed.Here's the tooth in occlusal view:
 Bison have a selenodont tooth pattern with two main crescent-shaped cusps, similar to many other artiodactyls. The tooth is worn, but only slightly, so this animal was likely a young adult when it died. The tooth had not yet worn down as far as the stylid on the lingual surface, so the enamel loop isn't visible; had the animal lived longer the loop would eventually have appeared as the tooth wore down.This tooth was found near the east dam of Diamond Valley Lake.
Bison have a selenodont tooth pattern with two main crescent-shaped cusps, similar to many other artiodactyls. The tooth is worn, but only slightly, so this animal was likely a young adult when it died. The tooth had not yet worn down as far as the stylid on the lingual surface, so the enamel loop isn't visible; had the animal lived longer the loop would eventually have appeared as the tooth wore down.This tooth was found near the east dam of Diamond Valley Lake.
Fossil Friday - peccary molar
 The Pleistocene megafauna from Diamond Valley Lake is dominated by a few prominent groups such as horses, bison, and mastodons. But there were a lot of species of mammals in the valley that are represented by only a limited number of fossil specimens. One of these rare types is the peccary, Platygonus.Peccaries, or javelinas, are artiodactyls that are sometimes referred to as "New World pigs". In fact, they are placed in their own family, the Tayassuidae, separate from domestic pigs and other Old World pigs (Family Suidae). Peccaries still live in Central and South America, and one species, the collared peccary, can still be found in the southwestern United States (the example below is at the Henry Doorly Zoo in Omaha):
The Pleistocene megafauna from Diamond Valley Lake is dominated by a few prominent groups such as horses, bison, and mastodons. But there were a lot of species of mammals in the valley that are represented by only a limited number of fossil specimens. One of these rare types is the peccary, Platygonus.Peccaries, or javelinas, are artiodactyls that are sometimes referred to as "New World pigs". In fact, they are placed in their own family, the Tayassuidae, separate from domestic pigs and other Old World pigs (Family Suidae). Peccaries still live in Central and South America, and one species, the collared peccary, can still be found in the southwestern United States (the example below is at the Henry Doorly Zoo in Omaha):
 Peccaries were much more diverse and widespread in North America during the Pleistocene, and several different genera are known from the fossil record. Various species of the extinct genus Platygonus have been found all over the United States (the skeleton below was on display at the Smithsonian Institution):
Peccaries were much more diverse and widespread in North America during the Pleistocene, and several different genera are known from the fossil record. Various species of the extinct genus Platygonus have been found all over the United States (the skeleton below was on display at the Smithsonian Institution):
 The Platygonus tooth shown at the top is an upper molar, probably the upper right first molar. It's only lightly worn, as is clear in lateral view:
The Platygonus tooth shown at the top is an upper molar, probably the upper right first molar. It's only lightly worn, as is clear in lateral view:
 This tooth was found near the east dam of Diamond Valley Lake, not far from where the museum is located.
This tooth was found near the east dam of Diamond Valley Lake, not far from where the museum is located.
Casting program underway
 When I was at the Virginia Museum of Natural History, one of our most successful endeavors was an active molding and casting program to produce replicas of fossils in the VMNH collection. After moving to the Western Science Center one of my top priorities was to start a comparable program here, and I'm happy to report that our casting efforts are now underway.Since the WSC staff is still learning how to make molds, we chose some Pleistocene horse teeth for our first molds. These teeth are relatively tough and they have a pretty simple overall shape, so they stand up to molding well. After sealing cracks in the tooth and attaching it to a clay base (above), we built a box (made of either clay or, in this case, Legos) around the specimen:
When I was at the Virginia Museum of Natural History, one of our most successful endeavors was an active molding and casting program to produce replicas of fossils in the VMNH collection. After moving to the Western Science Center one of my top priorities was to start a comparable program here, and I'm happy to report that our casting efforts are now underway.Since the WSC staff is still learning how to make molds, we chose some Pleistocene horse teeth for our first molds. These teeth are relatively tough and they have a pretty simple overall shape, so they stand up to molding well. After sealing cracks in the tooth and attaching it to a clay base (above), we built a box (made of either clay or, in this case, Legos) around the specimen:
 Once the box is built and sealed, we can pour liquid silicone over the specimen (being done here by our Collections Manager, Darla Radford):
Once the box is built and sealed, we can pour liquid silicone over the specimen (being done here by our Collections Manager, Darla Radford):
 Once the silicone is cured, the fossil can be removed from the mold, and liquid resin can be mixed and poured in:
Once the silicone is cured, the fossil can be removed from the mold, and liquid resin can be mixed and poured in:
 When the resin is cured, the resulting cast can be removed (two different horse teeth shown here):
When the resin is cured, the resulting cast can be removed (two different horse teeth shown here):
 At this point the cast is ready for painting, to resemble the original fossil:
At this point the cast is ready for painting, to resemble the original fossil:
 Establishing a casting program will greatly expand the things WSC can do with its collections in terms of exhibits and educational programs. Some of these hand-painted replicas will also be available in the museum's gift shop.
Establishing a casting program will greatly expand the things WSC can do with its collections in terms of exhibits and educational programs. Some of these hand-painted replicas will also be available in the museum's gift shop.
Fossil Friday - horse toe
 Some of the most common animals in the Pleistocene deposits around Diamond Valley Lake are horses. While we don't have any complete individual horse skeleton in the WSC collections, across all our different specimens we probably have just about every bone in the skeleton represented. Shown here is an isolated horse hoof.Horses have highly specialized feet, with only a single toe on each foot. That toe is Digit III, equivalent to our middle fingers (on the front feet) and our middle toes (on the back feet). Modern horses actually also have non-functional remnants of the 2nd and 4th toes, a relict of their multi-toed ancestors.Fingers and toes are made up of a series of bones called phalanges (singular: phalanx) that articulate end-to-end with each other; the articulations are the joints in our fingers and toes. In horses, the last phalanx, or ungal, is enlarged into an arc-shaped wedge of bone to support the weight of the animal. In life, the ungal is covered with a keratinized sheath, the hoof itself, which doesn't usually preserve in fossils.The image at the top shows a horse ungal in dorsal view, with anterior at the top. The scalloped area at the back of the bone is the articulation with the next phalanx in the toe.Here' same ventral view of the same bone:
Some of the most common animals in the Pleistocene deposits around Diamond Valley Lake are horses. While we don't have any complete individual horse skeleton in the WSC collections, across all our different specimens we probably have just about every bone in the skeleton represented. Shown here is an isolated horse hoof.Horses have highly specialized feet, with only a single toe on each foot. That toe is Digit III, equivalent to our middle fingers (on the front feet) and our middle toes (on the back feet). Modern horses actually also have non-functional remnants of the 2nd and 4th toes, a relict of their multi-toed ancestors.Fingers and toes are made up of a series of bones called phalanges (singular: phalanx) that articulate end-to-end with each other; the articulations are the joints in our fingers and toes. In horses, the last phalanx, or ungal, is enlarged into an arc-shaped wedge of bone to support the weight of the animal. In life, the ungal is covered with a keratinized sheath, the hoof itself, which doesn't usually preserve in fossils.The image at the top shows a horse ungal in dorsal view, with anterior at the top. The scalloped area at the back of the bone is the articulation with the next phalanx in the toe.Here' same ventral view of the same bone:
 The lateral view, showing the wedge shape (the articulation with the next phalanx is at the upper right):
The lateral view, showing the wedge shape (the articulation with the next phalanx is at the upper right):
 Note the how symmetrical this bone is in dorsal and ventral view. Horses are the only hoofed mammals in the valley that have a single toe, and as such are the only ones with a more-or-less symmetrical hoof-shaped ungal. Other hoofed mammals from these deposits, such as bison, have two toes with much more asymmetrical ungals.
Note the how symmetrical this bone is in dorsal and ventral view. Horses are the only hoofed mammals in the valley that have a single toe, and as such are the only ones with a more-or-less symmetrical hoof-shaped ungal. Other hoofed mammals from these deposits, such as bison, have two toes with much more asymmetrical ungals.
Fossil Friday - turkey
 It's the day after Thanksgiving in the US, so for today's Fossil Friday we have a probable turkey bone from south of Diamond Valley Lake.I found this bone by accident last week while looking for good examples of packrat teeth. While this fragment isn't much to look at, we have almost no other fossil turkeys in the WSC collection.This fragment is part of the distal end of the tarsometatarsus. This is a structure unique to birds and certain other dinosaurs, in which the ankle bones (the tarsals) and the foot bones (the metatarsals) are fused into a single unit. The proximal end articulates with the shin bone (tibiotarsus in birds), while the distal end has structures called trochlea that articulate with the toes. Most birds have three toes, and so have three trochlea at the end of the tarsometatarsus, as can be seen in the examples from modern turkeys shown below (from Thornton et al. 2012):
It's the day after Thanksgiving in the US, so for today's Fossil Friday we have a probable turkey bone from south of Diamond Valley Lake.I found this bone by accident last week while looking for good examples of packrat teeth. While this fragment isn't much to look at, we have almost no other fossil turkeys in the WSC collection.This fragment is part of the distal end of the tarsometatarsus. This is a structure unique to birds and certain other dinosaurs, in which the ankle bones (the tarsals) and the foot bones (the metatarsals) are fused into a single unit. The proximal end articulates with the shin bone (tibiotarsus in birds), while the distal end has structures called trochlea that articulate with the toes. Most birds have three toes, and so have three trochlea at the end of the tarsometatarsus, as can be seen in the examples from modern turkeys shown below (from Thornton et al. 2012):
 The WSC fragment is one of the trochlea, I think for either the second or third digit (the inside or middle toe). I'll need to compare it to a modern turkey bone to confirm which one, and unfortunately the turkeys you buy at the grocery store don't typically include the tarsometatarsus!Judging by their near-absence in our collection, it would seem that turkeys were relatively rare in the valley during the Pleistocene. This is a little surprising since vast numbers of turkey bones have been recovered from Rancho la Brea, which is only about 80 miles away from Hemet (more than 11,000 turkey bones have been found there, according to Bocheński and Campbell, 2006). The Rancho la Brea turkey was long considered to be its own genus, Parapava, but was reassigned to the same genus as the extant turkey as Meleagris californica by Steadman (1980). (Below is a reconstructed skeleton of M. californica on display at the George C. Page Museum.)
The WSC fragment is one of the trochlea, I think for either the second or third digit (the inside or middle toe). I'll need to compare it to a modern turkey bone to confirm which one, and unfortunately the turkeys you buy at the grocery store don't typically include the tarsometatarsus!Judging by their near-absence in our collection, it would seem that turkeys were relatively rare in the valley during the Pleistocene. This is a little surprising since vast numbers of turkey bones have been recovered from Rancho la Brea, which is only about 80 miles away from Hemet (more than 11,000 turkey bones have been found there, according to Bocheński and Campbell, 2006). The Rancho la Brea turkey was long considered to be its own genus, Parapava, but was reassigned to the same genus as the extant turkey as Meleagris californica by Steadman (1980). (Below is a reconstructed skeleton of M. californica on display at the George C. Page Museum.)
 It's possible that turkeys may have breeding at Rancho la Brea, and there are reasons to think that they may have been particularly susceptible to being trapped in tar (Bocheński and Campbell, 2006). It's also not clear that the turkey in the WSC collection is M. californica; it could be the modern wild turkey M. gallopavo or some other species. Even so, it's remarkable that turkeys were seemingly quite rare such a short distance away from Rancho la Brea, and may have interesting implications for different environmental conditions across California during the Pleistocene.References:Bocheński, Z. M. and K. E. Campbell, Jr., 2006. The extinct California turkey, Meleagris californica, from Rancho la Brea: Comparative osteology and systematics. Contributions in Science, Natural History Museum of Los Angeles County, No. 509:1-92.Steadman, D. W., 1980. A review of the osteology and paleontology of turkeys (Aves: Meleagridinae). Contributions in Science, Natural History Museum of Los Angeles County, No. 330:131-207.Thornton, E. K., K. F. Emery, D. W. Steadman, C. Speller, R. Matheny, and D. Yang, 2012. Earliest Mexican Turkeys (Meleagris gallopavo) in the Maya Region: Implications for Pre-Hispanic Animal Trade and the Timing of Turkey Domestication. PLoS ONE 7(8): e42630. doi:10.1371/journal.pone.0042630
It's possible that turkeys may have breeding at Rancho la Brea, and there are reasons to think that they may have been particularly susceptible to being trapped in tar (Bocheński and Campbell, 2006). It's also not clear that the turkey in the WSC collection is M. californica; it could be the modern wild turkey M. gallopavo or some other species. Even so, it's remarkable that turkeys were seemingly quite rare such a short distance away from Rancho la Brea, and may have interesting implications for different environmental conditions across California during the Pleistocene.References:Bocheński, Z. M. and K. E. Campbell, Jr., 2006. The extinct California turkey, Meleagris californica, from Rancho la Brea: Comparative osteology and systematics. Contributions in Science, Natural History Museum of Los Angeles County, No. 509:1-92.Steadman, D. W., 1980. A review of the osteology and paleontology of turkeys (Aves: Meleagridinae). Contributions in Science, Natural History Museum of Los Angeles County, No. 330:131-207.Thornton, E. K., K. F. Emery, D. W. Steadman, C. Speller, R. Matheny, and D. Yang, 2012. Earliest Mexican Turkeys (Meleagris gallopavo) in the Maya Region: Implications for Pre-Hispanic Animal Trade and the Timing of Turkey Domestication. PLoS ONE 7(8): e42630. doi:10.1371/journal.pone.0042630